===== ASTRONAUTICAL EVOLUTION =====
Issue 67, 1 March 2011 – 42nd Apollo Anniversary Year
1. Does evolution progress? The multi-shell model of life shows how.
All content is by Stephen Ashworth, Oxford, UK,
unless attributed to a different signed author.
=============== AE ===============
(1) Does evolution progress? The multi-shell model of life shows how.
An article by a pair of US sociologists published recently by the left-wing Monthly Review (vol.62, no.9, Feb. 2011) serves as a reminder that the concept of progress, while central to the astronautical endeavour, still arouses impassioned debate.
“Stephen Jay Gould’s Critique of Progress” by Richard York and Brett Clark asks to what extent the evolution of life on Earth and the development of human society have unfolded in a deterministic manner, or alternatively to what extent they have been shaped by chance events. If the one tendency is predominant, then the appearance of the particular forms of life and society that we actually find in the biological and historical records is in some sense predetermined, while the effect of the other tendency would be that life and society could have turned out very differently.
Are life and society – we might ask – more like a flower, which emerges predictably through organised cellular division from seed, to bud, to bloom, to new seed? Or are they more like a cloud, which is blown unpredictably this way and that by the four winds of heaven, randomly changing direction and shape, dividing and recombining, before it finally evaporates?
Or maybe society is fundamentally different from biology? Yet society is still a living system, an outgrowth and an intimate part of a wider biota, and comparisons with that wider biology seem inevitable.
Stephen Jay Gould was firmly opposed to the notion of progressive biological evolution, whereas other biologists such as Richard Dawkins and Simon Conway Morris have defended the concept. All three writers have published books on evolution which are accessible to a popular audience, and all three are articulate authors who are well worth reading.
The concept of progress, argue York and Clark, results in the common presumption “that the emergence of Homo sapiens is the inevitable apex of evolutionary processes”. But those who argue in this way “make a common error in equating what we value most in our world with what is necessary or inevitable”, conclude the authors in the final paragraphs of their article. “The belief in progress is a prime example of how social biases can distort science. Gould aimed to show that the natural world does not conform to human aspirations. Nature does not have human meaning embedded in it, and it does not provide direction to how humans should live. We live, instead, in a world that only has meaning of our own making.” The result is, Gould stressed, “that we make our own history and that the future is open”. This conclusion is “similar to Karl Marx and other radical thinkers” – clearly a very different Karl Marx to the one who claimed that the social conditions of capitalism would inevitably lead to communism. (*)
[ (*) “Karl Marx saw the British experience as an expression of historical logic. Capitalist production had its laws: ‘It is a question of these laws themselves, of these tendencies working with iron necessity towards inevitable results. The country that is more developed industrially only shows to the less developed, the image of its own future.’ ” – David Landes, The Wealth and Poverty of Nations, Abacus, 1998, p.236.]
Without offering a book-length response in this posting, I should nevertheless like to make some basic points.
* * *
(1) York and Clark clearly believe (along with Gould) that a scientific view of the world refutes the idea of progress. This is factually incorrect. Consider the career of life on Earth as revealed by two centuries of paleontology:
- When Earth first formed, it had no life at all;
- For about 2 billion years, starting from 3.8 billion years ago, it had life, but only simple, procaryotic, bacterial life;
- For the next billion years or so it had, in addition to bacterial life, complex eucaryotic single-celled life;
- Subsequently, multicellular life began to form, and for the most recent past half billion years life on Earth has included bacterial, eucaryotic and multicellular forms.
What are we to call this process? The picture is one of the progressive accumulation of complex organisation in life. Progress in biology on the broadest scale is a well-documented fact.
* * *
(2) So why does Gould (together with his sociological admirers) deny this fact? One element in Gould’s view is that he regards complexity as being on a continuous scale. Organisms started simple, Gould says, so the random walk of evolutionary change is bound to throw up more complex organisms as time goes on. This is not because of a pre-existing direction built into evolution, but merely the result of random changes filling up the space of possibilities from one end, that of the greatest simplicity.
In fact, as I have already indicated, on the broadest scale complexity is not continuous, but rather hierarchical. Or if you prefer, the continuity of increasing complexity is split over several different levels or domains of life.
There is a space of possibilities for simple bacterial organisms, which they fill up over time in the way that Gould described, purely haphazardly. There is another space of possibilities for eucaryotic cells (which are more complex organisms, combining their bacterial precursors into nuclei, mitochondria and other units within the cell). But – and this is the key point – the range of possibilities for bacteria does not just merge gradually into that for eucaryotic cells. Instead there is a threshold which must be crossed, a precise development that creates the first eucaryotic cell. This is a bottleneck in evolution. Similarly, there is a threshold development which creates the first multicellular organism out of eucaryotic cells.
* * *
(3) What we have, therefore, drawn from the best available scientific picture of the past, is a system in which the landscape of possibilities is divided into specific domains connected by narrow gateways. When life gets started it is at a bacterial level. After billions of years of bacteria going this way and that under the pressures of change, one bacterial species stumbles haphazardly across the threshold leading to the eucaryotic domain.
If it makes a successful crossing of the threshold, it radiates a new variety of eucaryotic species. Ultimately one of these stumbles across the threshold of multicellular life, and again a successful crossing of that threshold leads to a new radiation, this time of multicellular forms. Experience with life on Earth has shown that each new domain, if established successfully, may thereafter persist over geological time.
A good image for this process on a particular planet would be a set of nested shells, each shell representing the space of possibilities for life on a particular level or domain, shown as circles in the diagram below. Let us call this the MULTI-SHELL MODEL of life (or, if you like, the multi-shellular model):
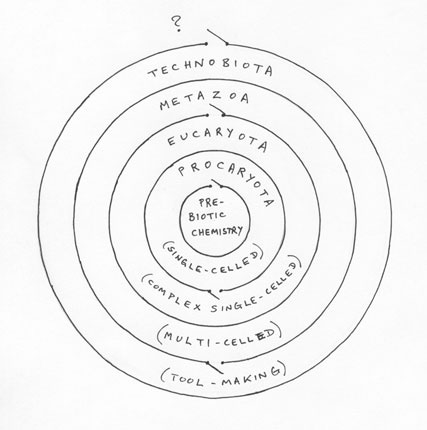
The innermost space represents the pre-biotic organic chemistry from which the procaryota, the simplest bacterial cells, emerge, in the next shell. One may imagine a number of balls bouncing around in each shell, modelling species (chemical, then biological) changing over time. To begin with, only the innermost circle is populated, representing conditions on Earth some 4 billion years ago, but over time a ball occasionally bounces through the first gateway and begins to populate the next shell.
Each ball thus moves from one point to another in a random fashion, bouncing off the walls, from time to time vanishing (as a species becomes extinct) and from time to time splitting into two (as a species generates multiple successor species). Under clement environmental conditions, more new species will appear than go extinct, until a domain is filled with some maximum population of different species.
The shells are nested for two reasons. As life becomes more complex, so the range of different possibilities available to it becomes greater – there is a greater variety of permutations of multicellular life than of single-celled life, so the multicellular domain takes up more space in the diagram. And secondly, a world containing life at one level also contains life at all the earlier levels – in our present-day situation the multicellular life of which we humans are a part coexists with single-celled life, and in fact could not exist without it. Thus the different levels of life are like storeys in a building: one may have a one-storey building, a two-storey or a three-storey building, but however high the topmost floor is, all the lower floors must be in place to support it, and the lower floors must have been built before the higher ones.
The narrow gateways that exist between one domain and another make the diagram resemble a labyrinth puzzle. As balls (species) bounce around in the domain of the procaryota, then, a small chance exists that one will randomly find the gateway into the next domain up, and the more balls there are and the longer they have been in existence, the greater the likelihood that one of them will happen to pass through this gateway.
Under clement environmental conditions, the first ball that passes into a vacant domain will produce offspring before it goes extinct, and thus will eventually fully populate that new domain.
Clearly, on this model progress within a single domain is not particularly meaningful. A species grows legs, a later species returns to the sea and loses those legs again (ichthyosaurs, dolphins); a species develops eyes, a descendant species makes a living underground and loses those eyes (a type of salamander); maybe a species develops a large brain, but then creates a utopian society where nobody has to think for themselves any longer, and its brain size shrinks again (socialism?). Progress on the multi-shell model is generated entirely by the ratchet effect when species cross the threshold into a new domain, which can only happen by chance, not by design or intent.
Some caveats. Clearly, if environmental conditions in one domain are inclement, then its population will tend to be driven towards zero species. If the more complex life is, the more dependent it is upon narrow ranges of temperature, atmospheric composition, soil salinity and so on, deteriorating environmental conditions on a planet will drive the higher domains of life extinct first. Large mammals (such as humans) will go extinct before algae do. Single-celled archaea at the ocean bottoms are almost immune to extinction on any timescale of less than tens of billions of years.
Again, while all the gateways are shown open, on other planets a particular gateway may be closed. On a world like Europa, for example, in which life (if it exists) is confined to a subsurface ocean, there may be simply no way for multicellular life to form, given that photosynthesis of oxygen is impossible.
A third caveat is that the diagram is based on terrestrial experience. On other planets other domains of life may be possible that are not found here, or a different geometry of relationships between domains. One can only work from what one knows, and meanwhile continue to explore other worlds.
But the essential take-away points from the multi-shell model are that life as we know it is arrayed on a hierarchical structure of qualitatively different domains, and this underlying structure defines a direction of progress.
* * *
(4) Let us return to the question of why Gould resisted the idea of evolutionary progress. One could hardly suppose that a great biologist like Gould imagined that multicellular creatures amounted to no more than unusually large and complex bacteria!
Surely the answer is that if one talks of progress in evolution, there creeps in a sense of some mystical or supernatural force – some godlike figure behind the scenes nudging the system forward with divine guidance. But evolutionary biology is founded on the axiom that change at any particular moment is fully accounted for by all the local conditions obtaining at that moment. There is no intrinsic trend towards accumulating greater complexity in organisms, no drive towards intelligence, and no way for the future to reach back and cause changes in the past. And there is absolutely no hidden intelligence manipulating life towards a pre-conceived outcome.
So we have a dilemma: over the long term, life certainly does progress in some sense (otherwise humans could never have evolved), but there is no plausible mechanism to define the direction of progress or to nudge evolutionary changes in that direction. One seems to be trapped between two opposing solutions, neither of which is particularly satisfactory:
- Our own existence is a random fluke thrown up by a blind nature; but then how do we account for the fact that the evolution of conscious intelligence is even possible at all? How can we account for the creative hierarchy of domains of life, if the only explanatory mechanism at our disposal comes down to atoms in motion under the power of random forces?
- Our own existence is the outcome of a universe pre-programmed to progress to successively more sophisticated domains of life; but then how do we account for the origin of that program? And how can it exert any influence over evolution, without bringing in fantasies of a divine hand or an angel materialising out of nowhere and offering a little divine protection to some herbivore that was about to be eaten, or a spot of manna from heaven to some carnivore that was about to starve, or tweaking the trajectory of some asteroid or the temperature of some volcano so that it does or does not cause the extinction of some species at some specific time?
Ultimately, it comes down to which of two impossible things one finds easier to believe:
- Whether humans have somehow evolved anyhow, despite there being no ghost of an explanation as to how it is even possible for such complex thinking organisms to exist at all in an apparently incommensurable universe of unconscious, inanimate mass-energy and spacetime;
- Or whether humans are part of some broader pattern of progressive evolution from an inanimate universe to a living one, despite there being no ghost of an explanation as to where that plan came from or how it is implemented in practice.
My claim here is that the multi-shell model of life goes a long way towards resolving the dilemma.
The model incorporates both random change and directed change. Within any one shell, one domain of life, change is random and subject entirely to local forces alone. Within the shell, the Gouldian paradigm is secure: there is change, but no overall progress to any long-range goal beyond the immediate survival and procreation of each organism.
But the overall, multi-shell, landscape of possibilities sets up a meaningful direction of change, and offers a mechanism through which random movements are converted to long-term progress in the direction of creating higher domains of life. It will not do so on all terrestrial planets, but it will do so on some, while the gateways to higher domains remain open.
The origin of the particular geometry of possibilities represented by the multi-shell diagram remains mysterious, obviously, but then so does the origin of the possibility that humans can exist at all in an overwhelmingly inanimate, unconscious universe. At least by recognising that the underlying structure of the universe is friendly to life in all its manifestations, we incorporate our own existence into our broadest picture of the universe. We are left with a puzzle as to why this should be so, but this is at least no worse than the puzzle of why it should have been any other way.
The multi-shell model removes any need for a supposed mystical urge or guiding hand towards the evolution of more complex forms of life by confining that mystical or invisible or guiding element to the equally invisible abstract concept of the landscape, or design space, of all possible life-forms. It is the hierarchical structure of this highly theoretical concept, with narrow gateways from one shell to the next, which makes evolutionary progress meaningful in the real world.
* * *
(5) Let us finally look to human society and to the future of evolution. The reader will have noticed that I have labelled the outermost shell “technobiota” (though further possibilities beyond even that are left open with a suggestive question mark). Here a few more reasonably securely known facts will help:
- Almost all the matter and energy of our Solar System is not claimed by living creatures for their sustenance, a situation clearly prevalent across the Galaxy;
- A very much larger quantity of the matter and energy in our own and other planetary systems than is available on Earth alone could be used to support much larger and more varied populations of living creatures through the use of space technologies and other technologies, hence the definition of the next great diversification of life as “technobiota”.
That diversification has not yet happened, in our own or in any neighbouring galaxy, so far as it is possible to tell. But were it to take place, the species which stumbled onto this new evolutionary radiation in this new environment would surely look very much like our own at this present juncture in history, leading one to conclude that Homo sapiens actually finds itself at present right in the middle of the gateway between the metazoan and technobiotic domains of life.
The key point about our species is not that we use tools (many species will pick up a stick or a stone and use it as a tool), nor even that we make tools (chimpanzees have been known to fashion a stick to the length required for a particular task). The point is that we use tools to make tools, in other words we systematically and routinely turn our tools back onto themselves in order to make better tools, starting with the basic stone-age technology of using one stone to put a cutting edge on another stone. This revolution in tool-making is still in full swing, and its final form is impossible to predict. The tools are taking on a life of their own, just as, half a billion years ago, the multicellular animals took on a life of their own which transcended the activities of individual cells.
What is ironic about the views of Stephen Jay Gould and others who decry the notion of progress is that we are just at the stage at which the nature of our own progress should be most clearly visible.
This is not to say that very many people are likely to understand what evolutionary progress means, let alone that they should approve of it. After all, when blue-green algae began to transform Earth’s atmosphere by releasing free oxygen, they had neither the knowledge nor the intention that they were setting the planet up for the diversity of multicellular animals which needed oxygen to breathe.
So is evolution a random walk or a pre-programmed unfolding of hidden potentialities? Clearly it is both, thanks to the underlying structure of the possible. While people’s understanding and approval of our transition to the technobiotic domain are not strictly necessary for that evolutionary development to happen, that intellectual engagement would certainly make the transition more pleasant for all concerned.